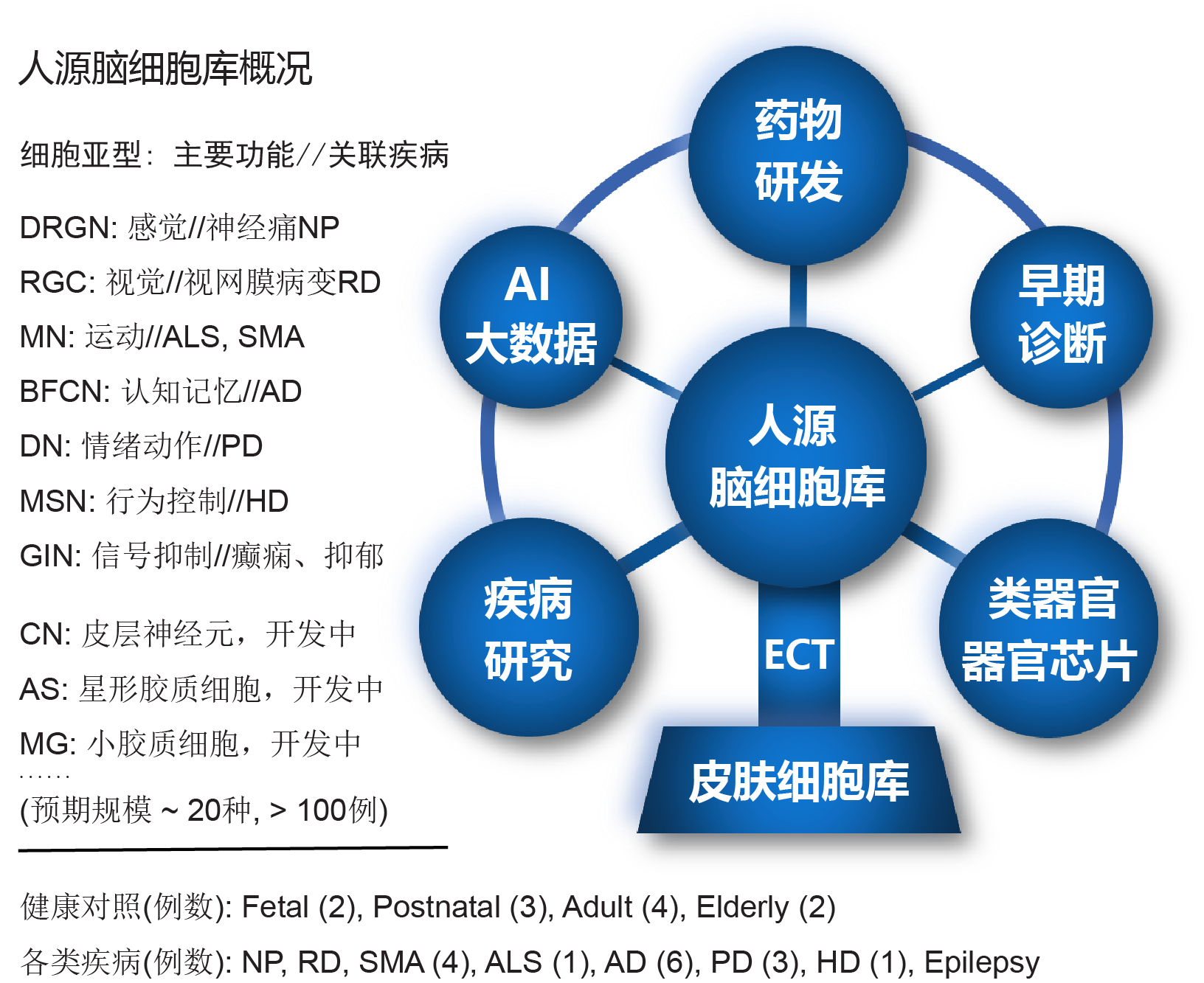
细胞产品
培养基/试剂
服务
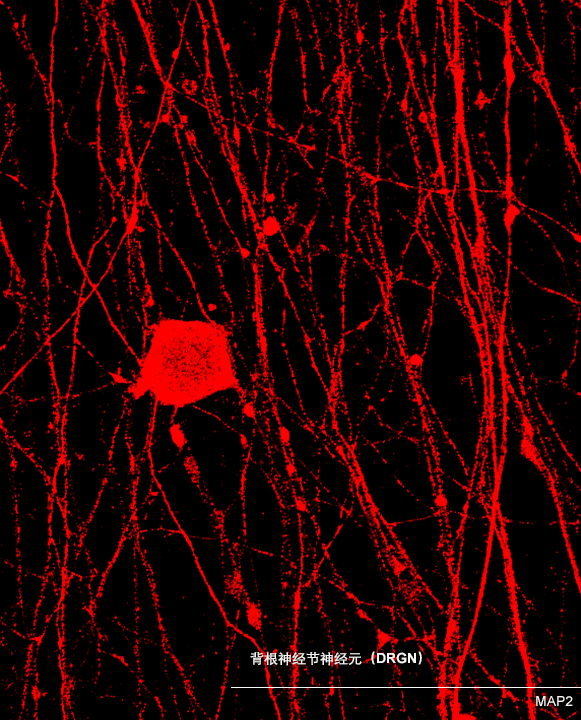
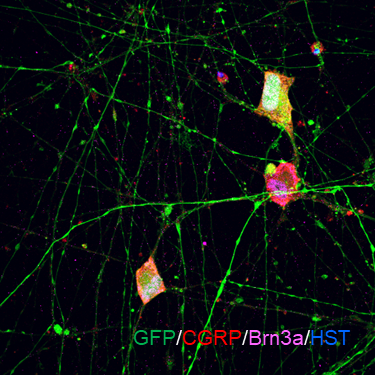
ECT-DRGN
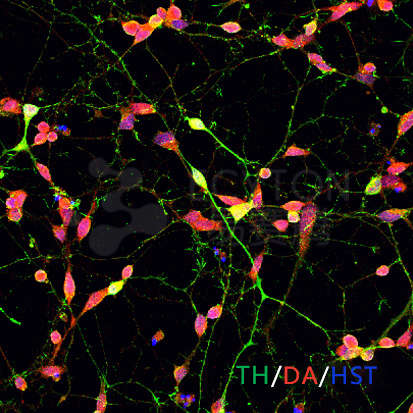
ECT-DN
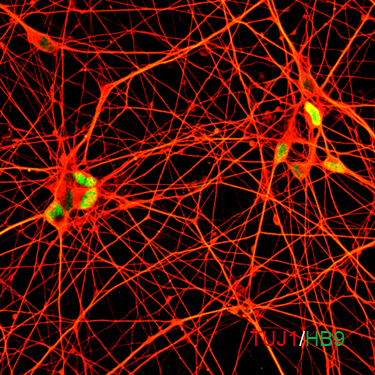
ECT-BFCN
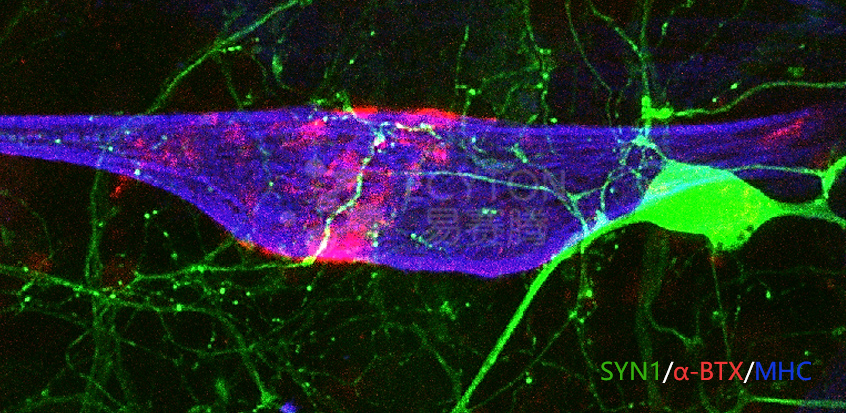
ECT-MN/Myotubes
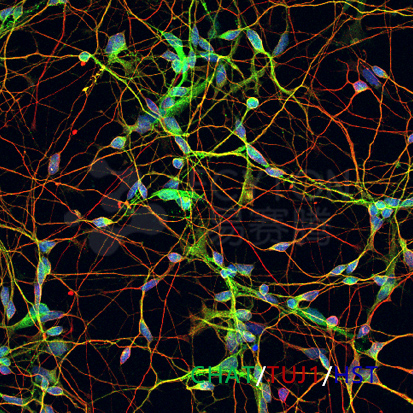
ECT-MN
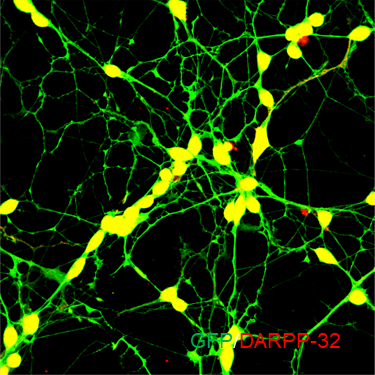
ECT-MSN
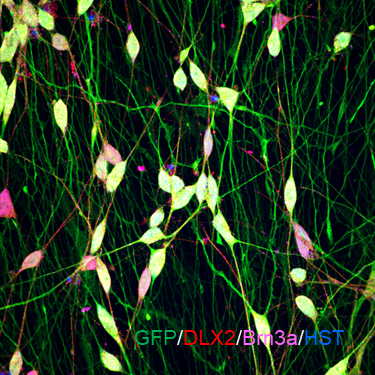
ECT-GIN
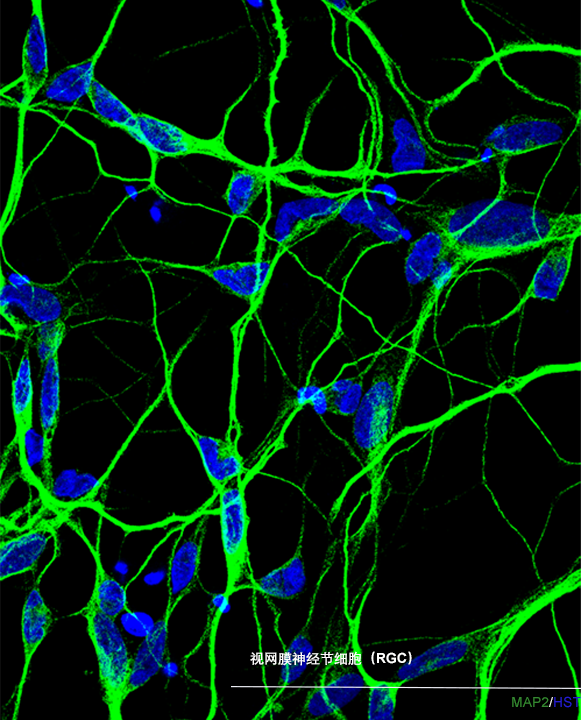
ECT-RGC
(DRGN601)
适应症:外周神经痛
(MNALS01)
(MNSMA01)
适应症:肌萎缩侧索硬化症
适应症:脊髓性肌萎缩症
(BFCNAD01)
适应症:阿尔茨海默病
我们的专家团队将及时回复您的问题